检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
cGAS-STING通路是负责识别胞质DNA免疫应答的主要通路 1。cGAS作为胞质DNA受体,可以被DNA和/或Mn2+ 激活并利用ATP和GTP合成第二信使2’3’-cGAMP,后者进一步激活STING并诱导I型干扰素等细胞因子的产生,从而介导抗感染/肿瘤免疫反应。研究表明多种病原微生物入侵及各种压力胁迫,如氧化应激、代谢紊乱及DNA损伤等都可导致胞质DNA的累积及Mn2+ 浓度的升高,从而激活cGAS-STING通路。因此,cGAS-STING信号通路在抵抗病原微生物感染、肿瘤及多种免疫相关疾病的发生及治疗中都发挥关键作用。
磷脂酰肌醇磷酸(phosphoinositides, PIPs)是构成真核生物细胞膜组分的重要磷脂(占总磷脂的5-10%),也是重要的信号分子 2。由于肌醇六元环上D-3,D-4或D-5位都可发生磷酸化修饰,因此真核生物中总共存在7种不同PIP分子。PI4P(phosphatydyinositol 4-phosphate)是胞内含量最高的PIP分子,广泛分布于各种膜组分,且在反式高尔基体(trans-Golgi network, TGN)上的含量最高。细胞中的PI4P的含量主要受其合成酶PI4Ks (PI4KA, PI4KB, PI4K2A和PI4K2B)和降解酶SAC1的调控。PI4P不仅是合成PIP2和PIP3的前体,也是重要的信号分子,在膜泡运输、脂质转运和细胞器形态维持等方面发挥关键作用。
2021年蒋争凡实验室利用STING自激活突变体诱导细胞死亡的特性,在HT080细胞中进行了基于CRISPR-Cas9介导的全基因组筛选,发现高尔基体腔内合成的硫酸化糖胺聚糖(sGAGs)可以通过结合到STING跨膜区的腔内侧,从而诱导STING的寡聚及激活 3。该论文的讨论部分提到STING和sGAGs的相互作用受到STING腔内侧pH值的调控,并由此推测STING在pH更低的后高尔基体囊泡(post-Golgi vesicles)中会有更高的活性。但是在这个研究中,他们没有解释STING为什么需要及如何转运到后高尔基体囊泡。
2023年3月14日,威尼斯wnsr666蒋争凡实验室在《Immunity》上以research article形式在线发表了“ARMH3-mediated recruitment of PI4KB directs Golgi-to-endosome trafficking and activation of the antiviral effector STING”,报道了ARMH3作为接头蛋白招募PI4KB到STING,PI4KB通过合成PI4P促进STING从高尔基体到内体的转运(Golgi-to-endosome trafficking)及维持STING激活所需要的脂膜环境(lipid environment),从而帮助STING更加稳定高效的激活。
本工作首先通过CRISPR-Cas9介导的全基因组筛选在HeLa细胞系中鉴定到一个未知功能的新基因C10ORF76 (又名ARMH3)对STING的激活十分重要(图一A)。ARMH3是一个鲜少被研究的蛋白,为数不多的几篇文章暗示了它对于PI4KB发挥激酶活性来磷酸化磷脂酰肌醇(phosphatidylinositol,PI)生成PI4P十分重要。免疫荧光实验发现STING激活后和ARMH3,PI4KB及PI4P都在反式高尔基体有明显的共定位。进一步免疫共沉淀实验显示,STING只有在ARMH3存在的情况下和PI4KB有互作(图一B),表明ARMH3在STING和PI4KB之间起到桥梁作用。进一步实验表明ARMH3分别结合位于STING第二、三跨膜区之间的连接区域及位于PI4KB激酶结构域中间的连接区域。突变掉STING负责结合ARMH3的氨基酸或者利用CRISPR-Cas9原位删除PI4KB与ARMH3互作的关键区域(6个氨基酸残基)都会显著削弱cGAS-STING通路的激活。这些结果表明ARMH3招募PI4KB对于STING的激活十分关键。更重要的是,细胞内敲除ARMH3或者原位删除PI4KB结合ARMH3所需的6个氨基酸,STING在上游通路活化后几乎呈现完全的内质网定位,暗示如果2’3-cGAMP结合后被转运到高尔基体的STING无法被继续转运到后高尔基体囊泡,将会被细胞内COPI/AP1介导的从高尔基体向内质网的逆向转运运回内质网,并完全抑制STING的活化。
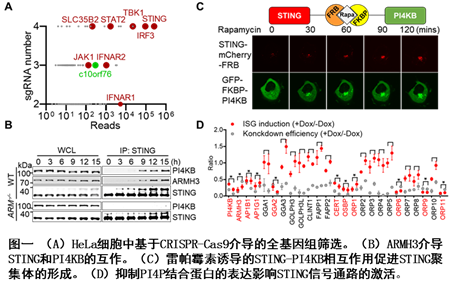
为了更直接的研究PI4KB对STING激活的影响,他们使用雷帕霉素诱导的FKBP和FRB相互作用,直接招募PI4KB-GFP-FKBP到STING-mCherry-FRB,并发现招募PI4KB到STING会显著促进STING聚集体的形成及下游信号通路的激活(图一C),且这种对于STING通路的激活能力完全依赖于PI4KB的激酶活性。相反,招募PI4P的磷酸酶SAC1到STING则会抑制STING聚集体的形成及下游信号通路的激活。表明PI4KB通过在STING周围合成PI4P促进STING的激活。有趣的是,SAC1的敲低会引起的细胞内PI4P水平的异常升高,进而导致STING不依赖于cGAS的自激活。TMEM39A是一个自身免疫性疾病相关基因且被报道参与细胞内的PI4P水平的调控 4。同样,TMEM39A的敲低也会导致细胞内PI4P的水平的异常升高及STING的自激活,该发现有助于理解TMEM39A相关自身免疫疾病的致病机制。此外,SCAP 和NPC1的突变也被报道可以引起细胞内PI4P的水平的异常升高及STING的自激活 5-8,提示胞内PI4P水平的异常升高可能是这些基因突变导致STING自激活的共同原因。
为了研究PI4P如何影响STING的激活,他们对23个PI4P结合蛋白进行了系统性地敲低并发现其中9个成员的降低表达会使STING的激活严重受损(图一D)。功能分析表明这9个PI4P结合蛋白可以分为两类。其中AP-1和GGA2负责介导高尔基体到内体的膜泡运输。由于内体中相对更低的pH值有利于STING与sGAGs的结合,他们推测PI4P 可以通过指导AP-1和GGA2介导的clathrin-coated vesicles (CCVs) 将STING从高尔基体转运到内体从而避免STING被运回内质网,并且更加强烈激活STING。另一类PI4P结合蛋白均为脂质转运蛋白,其中OSBP和ORPs负责从内质网向高尔基体或内体转运胆固醇,CERT负责神经酰胺从内质网到高尔基体的转运。神经酰胺在高尔基体上会在SGSM1的催化下转化为鞘磷脂,且SGSM1在细胞内的敲除会显著削弱STING的激活,提示鞘磷脂而非神经酰胺直接作用于STING。通过特异的化学小分子破坏胆固醇和鞘磷脂在脂膜上的分布会使STING聚集体逐渐消散并显著抑制STING下游信号通路的激活,表明胆固醇和鞘磷脂对STING聚集体的形成和激活十分关键。以上结果共同表明PI4P还可以通过脂质转运蛋白维持STING活化所需的脂膜环境进而影响STING的激活。此外,Lyz-Cre介导的Armh3条件敲除小鼠实验表明Armh3在机体对抗DNA病毒感染过程中发挥关键作用。
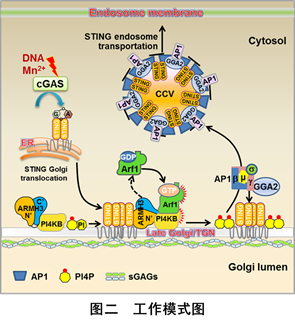
综上所述,该研究发现:1)STING为什么需要离开及如何离开高尔基体(后高尔基体转运(post-Golgi trafficking))的分子机制(图二);2)一个功能未知蛋白ARMH3(可能还具有GEF活性,激活膜泡转运中非常重要的Small GTPase Arf1)在cGAS-STING通路中的重要作用;3)脂膜环境对STING的活化十分关键;4)细胞内PI4P水平的异常升高可以导致非依赖于cGAS的STING分子自激活(cGAS-independent STING autoactivation)。重要的是,这种现象在很多脂质代谢紊乱的自身免疫病患者中被发现及报道。鉴于PI4P在脂质运输及脂质稳态维持中的核心作用,他们的研究还提示STING作为一个潜在的细胞脂质稳态感受器(lipid homeostasis sensor),可能是细胞内的一种新型危险信号感受器。
威尼斯wnsr666生科院博士后方润和蒋启飞为该文章的共同第一作者,2022级研究生贾新颖也为本研究做出贡献。生科院/北大-清华生命科学联合中心的蒋争凡教授为通讯作者。本研究工作得到了国家自然科学基金委、科技部国家重点基础研究项目、威尼斯wnsr666“细胞增殖与分化”教育部重点实验室及“北大-清华生命科学联合中心”的资助。
原文链接:https://doi.org/10.1016/j.immuni.2023.02.004
参考文献:
1 Fang, R. et al. Recent advances in the activation and regulation of the cGAS-STING pathway. Advances in Immunology 156, 55-102, doi:10.1016 /bs.ai.2022.09.003 (2022).
2 Posor, Y. et al. Phosphoinositides as membrane organizers. Nat Rev Mol Cell Biol, doi:10.1038/s41580-022-00490-x (2022).
3 Fang, R. et al. Golgi apparatus-synthesized sulfated glycosaminoglycans mediate polymerization and activation of the cGAMP sensor STING. Immunity 54, 962-975, doi:10.1016 /j.immuni.2021.03.011 (2021).
4 Miao, G. et al. The ER-Localized Transmembrane Protein TMEM39A/SUSR2 Regulates Autophagy by Controlling the Trafficking of the PtdIns(4)P Phosphatase SAC1. Mol Cell 77, 618-632 e615, doi:10.1016/j.molcel.2019.10.035 (2020).
5 Wakana, Y. et al. The ER cholesterol sensor SCAP promotes CARTS biogenesis at ER-Golgi membrane contact sites. J Cell Biol 220, doi:10.1083/jcb.202002150 (2021).
6 Kutchukian, C. et al. NPC1 regulates the distribution of phosphatidylinositol 4-kinases at Golgi and lysosomal membranes. EMBO J 40, e105990, doi:10.15252/embj.2020105990 (2021).
7 York, G. et al. Limiting Cholesterol Biosynthetic Flux Spontaneously Engages Type I IFN Signaling. Cell 163, 1716-1729, doi:10.1016/j.cell.2015.11.045 (2015).
8 Chu, T. et al. Tonic prime-boost of STING signalling mediates Niemann-Pick disease type C. Nature 596, 570-575, doi:10.1038/s41586-021-03762-2 (2021).
地址:北京市海淀区颐和园路5号
金光生命科学大楼
电话:010-62757794

北大生科官方微信

生声不息公众号